An Ecological Calculus: Estimating and Correcting Male Social Binding Deficits
The Contracted Ecology
The opening essay in this series asked about the measure of a man. The epidemiological data that framed the question — one in five young men in OECD nations reporting loneliness, three-quarters of global suicides accounted for by males, a doubling of the share of young American men neither in school nor employed — described the consequences of an ecological contraction, the biological mechanism of which was elaborated upon in the intervening essays. Thus, the measure that matters is not of the man but of his social ecology, which is, by any available metric, narrower than the one the species evolved within.
Default mode network (DMN) hyperconnectivity, associated with rumination across depression, anxiety, PTSD, and substance abuse (Hamilton et al., 2015; Berman et al., 2011), is a consistent neural signature of loneliness, with loneliness-linked differences in DMN-associated white matter tract integrity more prominent in men than in women (Spreng et al., 2020). A more complex social integration predicts survival with an odds ratio of 1.91, a figure that persists after controlling for age, sex, health behaviors, and baseline medical status (Holt-Lunstad et al., 2010). The multidimensional measures that produced this effect size — network diversity, group membership, frequency of contact — map onto several of the ecological dimensions this essay will develop.
The prevailing response to these phenomena has been to frame the problem in endocrine terms: men need testosterone for status-driven motivation, oxytocin for affiliative bonding, and cortisol regulation for stress buffering. In this framing, supplying the molecule could substitute for supplying the social condition that produces it. Hormones, however, are mediators in a coupled system, and altering one parameter without altering the system’s input produces unpredictable, sometimes paradoxical, results.
The sex-specificity of this ecological vulnerability is grounded in the preceding essays’ evidence. The cortisol-affiliation sex-crossover — rising cortisol after defeat predicts increased affiliation in women but decreased affiliation in men (Sherman et al., 2017) — means that ecological deprivation pushes men toward withdrawal while the same deprivation pushes women toward social reconnection. The DMN white-matter differences associated with loneliness are more pronounced in men (Spreng et al., 2020). The behavioral dimensions described in the Groups essay — affiliative withdrawal under stress, status-seeking through visible contribution, bonding through coordinated physical activity rather than conversation — predict that men’s wellbeing should be more tightly coupled to specific structural properties of social environments than to social contact per se. The calculus that follows is built on this sex-differentiated architecture.
The Calculus
Every environment places specific demands on the male’s processing capacity: demands for status inference, coalition categorization, motor synchronization with other bodies, norm enforcement, perspective-taking across asymmetric expertise, and goal-directed productive collaboration. Our neural processing systems evolved under a demand regime characterized by small-group, co-present, multi-sensory, temporally contingent interaction with high stakes and bounded membership. Contemporary ecologies deviate from this regime along specific, independently varying dimensions.
The calculus provides a means to interrogate a man’s weekly activities: what is the activity’s computational demand profile, and how does that profile compare to the demand regime under which the cognitive processing systems were calibrated? The difference between a brain simulating social life and a brain computing within social life — between mentalizing about relationships and participating in them — is the differential of interest. A man whose week includes competitive team sport, structured employment, and a long-standing friendship group may have adequate demand across most dimensions while remaining deficient in cross-experience scaffolding or normative accountability. A man who works remotely and socializes through gaming may have rich positional-differentiation demand alongside impoverished embodied-synchrony demand and no normative accountability at all.
Existing social-integration measures — the Berkman-Syme Social Network Index, the Lubben Social Network Scale, the UCLA Loneliness Scale — quantify contact frequency, network diversity, and subjective loneliness. The calculus departs from these instruments by indexing the type of computational demand an ecology supplies rather than the quantity of social contact it contains. A man with frequent social contact but no co-present physical coordination, no normative accountability, and no productive task orientation would score well on standard integration measures while the calculus would predict specific patterns of neural undersupply — and, accordingly, specific subjective signatures — that a contact-frequency measure would miss.
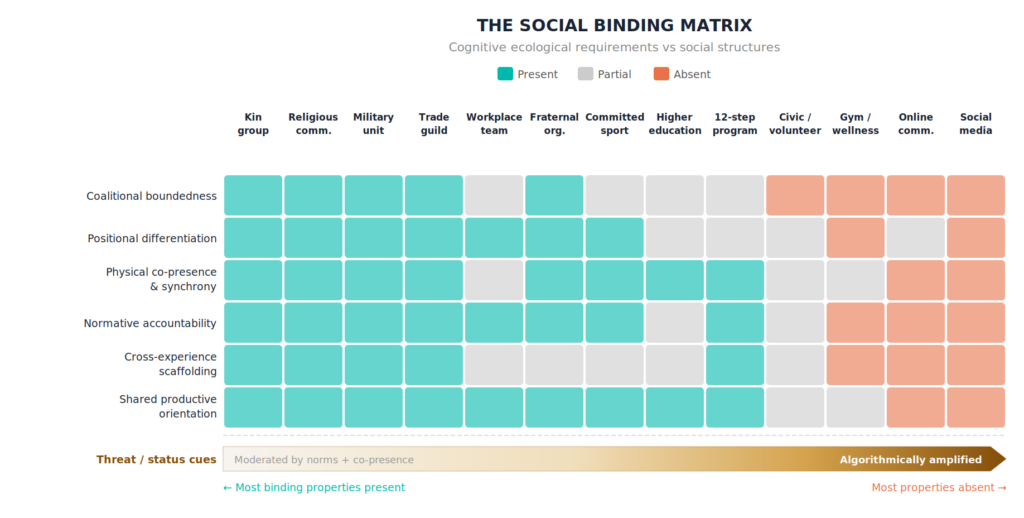
Six ecological dimensions organize the calculus. Each describes a property of the social environment rather than an individual trait, and each engages a separable component of the neural and endocrine systems that mediate male social behavior.
Ecological dimension — a property of the social environment, not an individual trait.
Computational demand — the processing load a social environment places on neural systems.
Decalibration — the drift in processing sensitivity that occurs when a neural subsystem is chronically undersupplied with its required input. The system does not idle; its parameters drift away from the range that would allow re-engagement with normal social input.
Recalibration — the restoration of processing sensitivity through sustained ecological input across the relevant dimensions.
Coupled system — a system in which neuromodulatory parameters and environmental inputs are mutually dependent. Altering one without altering the other produces unpredictable or paradoxical results.
The six ecological dimensions
Coalitional boundedness refers to the degree to which a group has defined membership, with costs attached to entry and exit, and includes members who are assigned or inherited rather than chosen for compatibility. The social categorization system — medial prefrontal cortex (mPFC) for self-group integration, fusiform face area for categorical face processing, temporoparietal junction (TPJ) for perspective-taking — tracks alliance rather than phenotype (Kurzban et al., 2001). The system evolved for partners who had to be made to work, not selected for agreeableness, and modern identity-based or interest-based grouping, where membership is voluntary and costless, may fail to engage the same coalitional circuits as functional, circumstantial coalitions. Costly entry stabilizes cooperation over time; costly exit raises the stakes of defection and makes the coalitional signal credible to the circuits that track it. A sports team with a roster, a religious congregation with attendance norms, and a workplace with an employment contract each supply this demand; open-access groups, drop-in events, and online communities with frictionless entry and exit do not.
Positional differentiation and contribution visibility refer to the degree to which members occupy recognizable positions within the group, contributions are visible to other members, and standing is subject to ongoing social feedback. The hippocampus and ventromedial prefrontal cortex (vmPFC) code linear rank structures (Kumaran et al., 2012); the mPFC mediates updating of one’s own position within that structure (Kumaran et al., 2016); the ventral striatum activates for outcomes with hierarchical consequences, paralleling its role in reward processing (Zink et al., 2008). Wingfield et al.’s (1990) challenge hypothesis linked testosterone rises during competitive status transitions as a context-dependent response to the ecology’s demand for status negotiation. Labor-force absence without a recognized role in any dependency structure produces near-uniform meaning-collapse in men, while labor-force absence in service of an outwardly recognizable caregiving role does not — consistent with a motivational system activated by a dependency structure in which the individual’s contribution is visible and consequential.
Physical co-presence and embodied synchrony refer to the degree to which interaction occurs in shared physical space and involves coordinated movement, synchronized activity, or tactile contact. The oxytocin system responds differentially to modalities of social contact. Salivary oxytocin measured during martial arts training showed that ground grappling (sustained close physical contact) produced significantly greater oxytocin increases than punch-kick sparring at equivalent heart rate and perceived exertion, isolating tactile contact as the active variable over exercise intensity (Rassovsky et al., 2019). Choral singing maintained oxytocin levels that solo singing did not (Schladt et al., 2017). Hyperscanning studies show inter-brain synchronization — dorsolateral prefrontal cortex (DLPFC) coupling during competitive interaction, TPJ coupling during cooperative interaction — during co-present coordinated activity (Lei et al., 2025; Zhang et al., 2023). The Spreng et al. (2020) neuroimaging data describe what happens when co-present social input is absent: the salience network is underactivated and the default mode network fills the gap with rumination. Digital interaction supplies partial input — the emerging hyperscanning literature shows measurable inter-brain synchrony during video-mediated and even text-based communication (Schwartz et al., 2024), though with fewer synchronized channels than co-present interaction.
Normative accountability refers to the degree to which cooperation within the group is sustained by mutual obligation and credible consequences for defection. Within-group prefrontal synchronization predicts intergroup conflict, and reduced DLPFC activity associates with stronger parochialism (Yang et al., 2020), meaning the prefrontal system’s role is, in part, to restrain in-group favoritism and enforce broader cooperative norms. Without norm enforcement, cooperation becomes risky — investing in a group without confidence in reciprocation exposes the individual to exploitation — and the executive control network suppresses affiliative investment in response. The social-support cortisol-buffering effect demonstrated by Heinrichs et al. (2003) required a best friend (a relationship with history, implicit norms, and mutual accountability), not a stranger assigned by a program. In turn, oxytocin supplementation studies have shown that activating the affiliative system in a context without normative structure produces amplified stress rather than buffering (Eckstein et al., 2014). A group that notices when a member is absent supplies a coalitional signal the categorization and affiliation circuits can register; a group that does not notice supplies ambient sociality rather than coalition.
The intervention-failure literature converges on the same prediction. In a systematic review of social-isolation interventions for older adults, group activities with an educational or shared-purpose component were consistently effective, while one-to-one befriending programs — which supply social contact without productive orientation, normative accountability, or coalitional boundedness — were not (Cattan et al., 2005). The calculus predicts this differential: befriending supplies contact quantity without the specific computational demands the processing systems require.
Cross-experience scaffolding refers to the degree to which the group embeds sustained relationships between members at different levels of experience, skill, or authority, in which the more experienced member models, corrects, and calibrates the less experienced member’s participation. The developmental calibration sequence traced through the preceding essays — from prenatal testosterone organizing receptor landscapes, through early paternal caregiving calibrating the oxytocin system, to prefrontal maturation bringing reactive aggression under cortical regulation — describes a series of calibration events, each requiring input from a more mature male system. The rearing father’s behavior, not his genotype, determined the son’s oxytocin receptor density and subsequent caregiving competence in the California mouse (Gleason & Marler, 2013). Primary-caregiving fathers activated the same parental brain network regardless of biological relatedness, with amygdala-STS connectivity scaling with time spent in direct caregiving (Abraham et al., 2014). The same experience-dependent neural activation, driven by behavior rather than biology, is the mechanism through which coaching, mentorship, and elder-junior pairing in group contexts can sustain calibration across the lifespan. A non-parental male authority figure providing structured physical-contact interaction partially re-engaged blunted oxytocin reactivity in at-risk adolescent boys (Harwood-Gross et al., 2020). Sustained complex social relationships of this kind drive experience-dependent structural adaptation, including increased gray matter in mid-STS and ventral insula (Sallet et al., 2011; Testard et al., 2022).
Shared productive orientation refers to the degree to which the group exists to accomplish, produce, or sustain something beyond the social contact itself. A group organized around a shared productive task simultaneously engages multiple neural systems: the task activates executive processing (Seeley et al., 2007), the collaborative structure activates self-position updating (Kumaran et al., 2016), and the output activates reward processing (Zink et al., 2008). A group organized around conversation or companionship alone engages the default mode network’s mentalizing circuits without recruiting the executive and reward systems. Men’s groups organized around building, repairing, cooking, training, or producing engage these circuits in ways that groups organized around conversation do not, because the affiliative circuits in men are more reliably engaged through shared physical activity and visible contribution than through conversation organized around emotional disclosure.
The coupling mechanism
Hormones are real-time parameters of the system’s operation, not downstream confirmation that the system has been engaged. Testosterone alters the gain of amygdala and orbitofrontal responses to status-relevant stimuli. Oxytocin shifts the salience weighting of social versus non-social cues in the anterior insula, primarily through GABAergic mechanisms that increase the signal-to-noise ratio (Froemke & Young, 2021). Cortisol modulates hippocampal memory consolidation for socially threatening events.
The system, therefore, has feedback structure. Ecological input drives processing engagement, which produces neuromodulatory change, which alters processing sensitivity, which changes what the organism detects and responds to in the next moment of ecological input. Testosterone rises during competitive status transitions and modulates subsequent competitive behavior, an effect that depends on both basal cortisol levels and environmental cues to opponent status (Knight et al., 2022; Inoue et al., 2023). Oxytocin released during synchronized co-present activity increases anterior insula sensitivity to subsequent social cues, lowering the activation threshold for the next round of social processing. The loop is self-reinforcing when the ecology supplies adequate demand across dimensions.
The converse is also self-reinforcing. When ecological input falls below the demand threshold for a processing subsystem, the neuromodulatory parameters that maintain that subsystem’s sensitivity begin to drift. Chronic social isolation in adolescent mice produces abnormally elevated oxytocin receptor levels in the prefrontal cortex (Li et al., 2026), downregulation of oxytocin receptor gene expression in the central amygdala contributing to anxiety-like phenotypes, and lasting changes in VTA dopaminergic neuron excitability that persist after regrouping (Musardo et al., 2022). OXTR DNA methylation status affects neural response to social stimuli in young adults: those with presumed decreased oxytocin sensitivity recruit compensatory attentional control networks to process social information that others process automatically (Puglia et al., 2023). Acute social isolation in humans evokes midbrain craving responses to social cues that parallel the craving responses produced by food deprivation, but chronically lonely individuals show blunted versions of this response (Tomova et al., 2020). The system does not simply idle under conditions of deprivation — it decalibrates, and decalibration makes re-engagement harder and non-linear, because the processing parameters are now less responsive to the inputs that would restore them.
| Ecological dimension | Ancestral reference | Contemporary deficit pattern | Neuromodulatory coupling | Decalibration signature |
|---|---|---|---|---|
| Coalitional boundedness | Kin/proximity groups with unambiguous membership determined by circumstance, not choice | Open-access communities, self-selected ideological groups, frictionless digital followings | Oxytocin promotes parochial prosociality within bounded ingroups (De Dreu et al., 2010); exit cost stabilizes cooperative commitment | Coalition-detection circuits run on ambient social media input; competitive-hierarchy activation without affiliative conversion |
| Positional differentiation &
Contribution visibility |
Small-group hierarchy with visible contribution, performance feedback, and positional consequences | Flat digital environments, remote work without feedback, egalitarian socializing without task differentiation | Testosterone modulates gain on amygdala/OFC status-evaluation circuits; competitive outcomes produce context-dependent T fluctuations (challenge hypothesis; Wingfield et al., 1990; Knight et al., 2022) | Reduced sensitivity to status-consequential input; subjective purposelessness even in the presence of social contact |
| Physical co-presence &
Embodied synchrony |
Co-present, multi-sensory, temporally contingent interaction involving coordinated movement and tactile contact | Digital-dominant social contact; sedentary socializing; conversation without physical coordination | Tactile contact during coordinated activity produces differential oxytocin release (Rassovsky et al., 2019); inter-brain TPJ/DLPFC coupling during co-present coordination (Lei et al., 2025) | DMN hyperconnectivity; increased rumination; social contact that produces mentalizing without embodied engagement |
| Normative accountability | Mutual obligation within kin/proximity groups; credible consequences for defection; reciprocal monitoring | Relationships without expectations; groups that do not notice absence; befriending programs without mutual obligation | DLPFC inhibitory function restrains parochialism and enforces cooperative norms (Yang et al., 2020); cortisol buffering requires a trusted relationship, not assigned contact (Heinrichs et al., 2003) | Affiliative investment suppressed as rational response to exploitation risk; the Eckstein et al. oxytocin paradox |
| Cross-experience scaffolding | Intergenerational mentorship; apprenticeship structures; sustained calibration relationships across experience levels | Age-segregated socializing; peer-only interaction; absence of sustained mentoring relationships | Rearing behavior determines offspring OTR density (Gleason & Marler, 2013); sustained complex social processing drives structural adaptation in mid-STS and ventral insula (Sallet et al., 2011; Testard et al., 2022) | Reduced gray matter in social-cognitive regions; absence of calibration relationships across the lifespan; slow-acting, cumulative deficit |
| Shared productive orientation | Groups organized around hunting, building, defense, provisioning — collaborative production with tangible output | Groups organized around conversation, companionship, or consumption without productive task | Task provides goal activating ECN (Seeley et al., 2007); collaborative structure activates mPFC self-position updating (Kumaran et al., 2016); output activates ventral striatum reward processing (Zink et al., 2008) | Executive and reward systems disengaged; DMN mentalizing without ECN recruitment; the neural profile Spreng et al. (2020) observed in lonely individuals |
Archetypal profiles
Four deficit patterns recur across the contemporary male ecology. Each corresponds to a specific combination of ecological dimensions that are undersupplied relative to the ancestral regime, and each produces a characteristic subjective signature. The profiles are described in terms of the six dimensions; in practice, most men present with compound deficits across several.
The man whose ecology is dominated by digital interaction and solo activity (Profile A) experiences deficits in co-presence/synchrony and normative accountability simultaneously. The salience network receives insufficient co-present social input to perform the DMN-to-ECN switch. The default mode network fills the processing gap with internally directed social simulation: mentalizing about relationships rather than participating in them. He may belong to online communities (partial coalitional boundedness) and hold a ranked position in a competitive game (partial positional differentiation), but these supply the categorization and status-tracking circuits without engaging the embodied synchrony or norm-enforcement circuits. The Spreng et al. (2020) UK Biobank data, drawing on approximately 40,000 participants, confirm that lonely individuals show greater within-network DMN functional connectivity and greater between-network DMN coupling with somatomotor and limbic regions. Physical activity partially compensates for this deficit, especially in individuals with the highest DMN connectivity (Benedyk et al., 2024).
The man without differentiated positions or status-consequential outcomes (Profile B) experiences a deficit in positional differentiation and contribution visibility. The vmPFC has no rank structure to code. The ventral striatum receives no hierarchically consequential outcomes to evaluate. The behavioral signature is purposelessness, meaning-collapse, the sense that one’s presence makes no difference. Testosterone modulates the sensitivity of the status-processing circuits, but it does not supply the input those circuits require. Increasing testosterone in this man increases the system’s reactivity to status signals without supplying the signals themselves, an intervention that the dual-hormone literature suggests may produce either status-seeking or status-loss avoidance depending on concurrent cortisol levels and environmental cues (Knight et al., 2022).
The man who has social contact without coordinated physical activity (Profile C) experiences a deficit in co-presence/synchrony while his other dimensions may be adequately supplied. He has identifiable groups, a job, and mutual obligations, but no activity in which participants’ actions are temporally coordinated and mutually contingent. His social life produces information exchange without inter-brain coupling. The computational demand profile predicts that coordinated physical activity — team sport, group singing, collaborative building — would engage the synchronization circuits that verbal interaction alone does not recruit.
The most prevalent contemporary male profile (Profile D) combines abundant positional-differentiation demand (workplace hierarchy, competitive gaming, social media ranking) with minimal co-presence/synchrony, weak normative accountability, no cross-experience scaffolding, and no shared productive orientation beyond employment. The status-processing circuits are loaded while the cooperative, synchronization, and calibration circuits idle. The behavioral consequence is competitive engagement that does not generate affiliative satisfaction: purpose without belonging. The modern ecology often supplies positional differentiation through contexts that lack all five other dimensions (remote work, solo competitive gaming), producing a man whose status-tracking system is highly active while everything else decalibrates. The calculus predicts that activities engaging the idle dimensions without displacing the active ones — a martial arts dojo, a volunteer fire company, a building crew, a long-running choral ensemble — would supply coalitional boundedness, co-presence with synchrony, normative accountability, cross-experience scaffolding, and shared productive orientation simultaneously without exacerbating competitive engagement, which is consistent with the evidence that activities of this kind produce the strongest intervention effects.
The manosphere ecology described in the opening essay maps onto a specific variant of Profile D: the status-tracking and coalition-categorization circuits are supplied by online ranking systems and ingroup/outgroup rhetoric, while co-presence, normative accountability, cross-experience scaffolding, and shared productive orientation are absent. The framework predicts that the behavioral output of this configuration — competitive intensity without affiliative satisfaction, dominance-seeking without coalitional bonding — is a predictable consequence of the demand profile, not a personality type.
| Profile | Primary dimension deficits | Dimensions adequately supplied | Subjective signature | Predicted recalibration target | Predicted recalibration latency |
|---|---|---|---|---|---|
| A: Digital-dominant ecology | Co-presence/synchrony; normative accountability | Partial coalitional boundedness (online); partial positional differentiation (gaming/work) | Disconnection despite active mentalizing about others | Structured co-present group activity with task demands and attendance norms | DMN-dominant processing may resist salience-network switching; salience sensitivity has drifted, predicting reduced reward response in early sessions |
| B: Positional deficit | Positional differentiation; contribution visibility | Variable — may retain coalitional structure and co-presence through existing social contact | Purposelessness despite social contact | Activity with defined roles, visible contribution, performance feedback, and consequential outcomes | Decalibrated status-processing circuits may initially fail to register hierarchical signals as rewarding; ventral striatum recalibration is gradual |
| C: Disembodied sociality | Co-presence/synchrony | Positional differentiation; coalitional boundedness; normative accountability | Social engagement without felt embodied connection | Coordinated physical activity — team sport, group singing, partner dance, collaborative construction | Transition from verbal-only to embodied interaction places initially high demands on an under-exercised synchronization system |
| D: Asymmetric loading | Co-presence/synchrony; normative accountability; cross-experience scaffolding; shared productive orientation | Positional differentiation (often heavily loaded) | Achievement without connection | Activities that engage all idle dimensions simultaneously — martial arts dojo, volunteer company, building crew, choral ensemble | Status-dominant ecology may suppress valuation of cooperative activities; recalibration requires sustained exposure before the reward system registers the new inputs |
The subjective signatures described above can serve as a rough self-assessment. Most men present with compound deficits across several profiles. The operative question is not whether a man is socially isolated but which dimensions of his ecology are undersupplied and what the specific pattern of undersupply predicts about his subjective experience.
Endocrine dysregulation as mismatch symptom
A man whose weekly ecology supplies all six dimensions does not need to attend to his endocrine profile. His testosterone fluctuations follow the competitive outcomes his ecology provides. His oxytocin release follows the synchronized co-present activity his ecology demands. His cortisol regulation follows the social buffering his accountable relationships supply. The hormones are responding correctly to an ecology that supplies what the processing systems evolved to require.
When that ecology erodes — when bounded groups, coordinated physical activity, normative accountability, and productive collaboration are replaced by digital, transactional, and solitary substitutes — the endocrine profile shifts accordingly. Testosterone fluctuations flatten because there are no competitive outcomes to fluctuate around. Oxytocin release attenuates because there is no synchronized co-present activity to trigger it. Cortisol regulation destabilizes because social buffering requires an accountable relationship to buffer within. These endocrine changes are the correct response to an impoverished ecology, not a pathology in themselves. In this context, supplementation treats the symptom while leaving the input deficit in place.
The plasticity window
Decalibration predicts that re-engagement will be harder than initial engagement. The processing parameters have drifted. The critical period literature offers evidence for this prediction — and a potential means of reversing it.
Nardou et al. (2019, 2023) demonstrated that oxytocin-mediated synaptic plasticity in the nucleus accumbens establishes a developmental critical period for social reward learning, and that psychedelics can reopen this window. A single dose of MDMA restored social conditioned place preference in adult mice — but only when administered in a social context. MDMA given to isolated mice produced the synaptic plasticity change (metaplastic upregulation of oxytocin-dependent long-term depression) without the behavioral change (restored social reward learning).
Psilocybin produces rapid dendritic spine growth in the frontal cortex — approximately 10% increases in spine density and head width within 24 hours of a single dose, persisting at least one month (Shao et al., 2021). In humans, psilocybin massively disrupts functional connectivity across cortex and subcortex, with some changes persisting for weeks (Siegel et al., 2024). These effects are sex-dependent in rodent models: testosterone increases 5-HT2A receptor density in male rat cortex (Sumner & Fink, 1998), and concurrent stress abolishes psilocybin’s post-acute behavioral effects in a sex-dependent manner (Farinha-Ferreira et al., 2025).
Pharmacological reopening of a plasticity window creates the potential for recalibration, but whether that potential translates to restored social processing in humans remains untested. The evidence to date is confined to rodent models for the critical-period mechanism (Nardou et al.) and to an observational study in humans that is consistent with the coupling prediction but cannot establish causality: across 1,200 participants at naturalistic gatherings, the mood-enhancing effects of psychedelic use were sequentially mediated by transformative experience and then by increased social connectedness (Forstmann et al., 2020). Social reconnection was part of the mechanism, not a side effect.
The coupled-systems framework generates a specific prediction: psychedelic-assisted recalibration of social processing should be most effective when paired with concurrent ecological restructuring — structured group activity that supplies the computational demands the reopened system needs to process. No trial has tested this prediction directly. The trial design that would test it would measure baseline social ecology across the six dimensions, administer the psychedelic with or without concurrent structured group activity, and track both neural and behavioral outcomes across the weeks during which the plasticity window remains open. Until such a trial is conducted, the prediction remains a derivation from the framework, not an established finding.
Conclusions
The argument developed across these essays traces a causal chain from genetics, through neuroendocrine pathways, to environment. The Y chromosome initiates a developmental cascade that organizes neural circuits for group life. Those circuits — the hierarchy-tracking system of the hippocampus and vmPFC, the coalition-categorization system of the mPFC and TPJ, the salience network that routes social input between DMN and executive processing, the synchronization circuits that couple co-present brains — are modulated by endocrine signals that are themselves calibrated by early caregiving and recalibrated by adult social context. The behavioral output — aggression type, affiliative stance, competitive-cooperative balance, coalitional categorization — is the product of this coupled system operating within a specific ecological demand regime.
Our social ecology has contracted, and the calculus developed in this parting essay offers a means to interrogate the contraction. Each of the six ecological dimensions engages a separable component of the neural and endocrine architecture described in the preceding essays. Where a dimension is adequately supplied, the corresponding subsystem operates within its calibrated range. Where a dimension is chronically undersupplied, the subsystem decalibrates — its parameters drift, its sensitivity decreases, and re-engagement becomes progressively harder. The endocrine markers that accompany this process — flattened testosterone fluctuation, attenuated oxytocin release, destabilized cortisol regulation — are the correct output of a system responding to impoverished input, not a pathology to be corrected pharmacologically. Recalibration follows from restoring the ecological input, and the endocrine profile follows the recalibration, not the reverse.
This model has limits. The near-complete absence of genetically informed research on human male affiliative behavior means that the heritability estimates for the dimensions most relevant to the calculus — coalitional bonding, synchronization sensitivity, normative accountability — do not exist. The critical-period plasticity mechanism, while demonstrated in rodent models, has not been tested in humans with the ecological-restructuring protocol the framework predicts would be necessary. The decalibration-recalibration dynamic, though consistent with the isolation and re-socialization literatures, has not been directly measured across the six dimensions in a prospective human cohort. The profiles and their predicted subjective signatures remain theoretical derivations pending empirical validation.
The measure of a man is not a set of individual attributes — not testosterone level, not dominance rank, not sexual conquest — but the fit between a man’s neural architecture and the social ecology within which he operates. Where the ecology supplies what his architecture evolved to process, the system functions. Where it does not, the system produces the withdrawal, the purposelessness, the rumination, and the dysregulated competition that the epidemiological data now document at population scale. The biology triangulates the cause, and the calculus triangulates the intervention: not in the man’s endocrine profile or personality type, but in the structure of the ecology his neural architecture still expects to find.
References
Abraham, E., Hendler, T., Shapira-Lichter, I., Kanat-Maymon, Y., Zagoory-Sharon, O., & Feldman, R. (2014). Father’s brain is sensitive to childcare experiences. Proceedings of the National Academy of Sciences, 111(27), 9792–9797. https://doi.org/10.1073/pnas.1402569111
Benedyk, A., Reichert, M., Giurgiu, M., Kossowski, B., Tost, H., Reif, A., Ebner-Priemer, U., & Meyer-Lindenberg, A. (2024). Real-life behavioural and neural circuit markers of physical activity as a compensatory mechanism for social isolation. Nature Mental Health, 2(3), 337–342. https://doi.org/10.1038/s44220-024-00204-6
Berman, M. G., Peltier, S., Nee, D. E., Kross, E., Deldin, P. J., & Jonides, J. (2011). Depression, rumination and the default network. Social Cognitive and Affective Neuroscience, 6(5), 548–555. https://doi.org/10.1093/scan/nsq080
Cattan, M., White, M., Bond, J., & Learmouth, A. (2005). Preventing social isolation and loneliness among older people: A systematic review of health promotion interventions. Ageing and Society, 25(1), 41–67. https://doi.org/10.1017/S0144686X04002594
De Dreu, C. K. W., Greer, L. L., Handgraaf, M. J. J., Shalvi, S., Van Kleef, G. A., Baas, M., Ten Velden, F. S., Van Dijk, E., & Feith, S. W. W. (2010). The neuropeptide oxytocin regulates parochial altruism in intergroup conflict among humans. Science, 328(5984), 1408–1411. https://doi.org/10.1126/science.1189047
Eckstein, M., Scheele, D., Weber, K., Stoffel-Wagner, B., Maier, W., & Hurlemann, R. (2014). Oxytocin facilitates the sensation of social stress. Human Brain Mapping, 35(9), 4741–4750. https://doi.org/10.1002/hbm.22508
Farinha-Ferreira, M., Miranda-Lourenço, C., Galipeau, C., Lenkei, Z., & Sebastião, A. M. (2025). Concurrent stress modulates the acute and post-acute effects of psilocybin in a sex-dependent manner. Neuropharmacology, 264, 110280. https://doi.org/10.1016/j.neuropharm.2024.110280
Forstmann, M., Yudkin, D. A., Prosser, A. M. B., Heller, S. M., & Crockett, M. J. (2020). Transformative experience and social connectedness mediate the mood-enhancing effects of psychedelic use in naturalistic settings. Proceedings of the National Academy of Sciences, 117(5), 2338–2346. https://doi.org/10.1073/pnas.1918477117
Froemke, R. C., & Young, L. J. (2021). Oxytocin, neural plasticity, and social behavior. Annual Review of Neuroscience, 44, 359–381. https://doi.org/10.1146/annurev-neuro-102320-102847
Gleason, E. D., & Marler, C. A. (2013). Non-genomic transmission of paternal behaviour between fathers and sons in the monogamous and biparental California mouse. Proceedings of the Royal Society B: Biological Sciences, 280(1763), 20130824. https://doi.org/10.1098/rspb.2013.0824
Hamilton, J. P., Farmer, M., Fogelman, P., & Gotlib, I. H. (2015). Depressive rumination, the default-mode network, and the dark matter of clinical neuroscience. Biological Psychiatry, 78(4), 224–230. https://doi.org/10.1016/j.biopsych.2015.02.020
Harwood-Gross, A., Feldman, R., Zagoory-Sharon, O., & Rassovsky, Y. (2020). Hormonal reactivity during martial arts practice among high-risk youths. Psychoneuroendocrinology, 121, 104806. https://doi.org/10.1016/j.psyneuen.2020.104806
Heinrichs, M., Baumgartner, T., Kirschbaum, C., & Ehlert, U. (2003). Social support and oxytocin interact to suppress cortisol and subjective responses to psychosocial stress. Biological Psychiatry, 54(12), 1389–1398. https://doi.org/10.1016/S0006-3223(03)00465-7
Holt-Lunstad, J., Smith, T. B., & Layton, J. B. (2010). Social relationships and mortality risk: A meta-analytic review. PLOS Medicine, 7(7), e1000316. https://doi.org/10.1371/journal.pmed.1000316
Inoue, Y., Burriss, R. P., Hasegawa, T., Yamagishi, T., & Kiyonari, T. (2023). Testosterone promotes dominance behaviors in the Ultimatum Game after players’ status increases. Scientific Reports, 13(1), 18029. https://doi.org/10.1038/s41598-023-45247-4
Knight, E. L., Morales, P. J., Christian, C. B., Prasad, S., Harbaugh, W. T., Mehta, P. H., & Mayr, U. (2022). The causal effect of testosterone on men’s competitive behavior is moderated by basal cortisol and cues to an opponent’s status. Journal of Personality and Social Psychology, 123(4), 893–911. https://doi.org/10.1037/pspa0000305
Kumaran, D., Melo, H. L., & Duzel, E. (2012). The emergence and representation of knowledge about social and nonsocial hierarchies. Neuron, 76(3), 653–666. https://doi.org/10.1016/j.neuron.2012.09.035
Kumaran, D., Banino, A., Blundell, C., Hassabis, D., & Dayan, P. (2016). Computations underlying social hierarchy learning: Distinct neural mechanisms for updating and representing self-relevant information. Neuron, 92(5), 1135–1147. https://doi.org/10.1016/j.neuron.2016.10.052
Kurzban, R., Tooby, J., & Cosmides, L. (2001). Can race be erased? Coalitional computation and social categorization. Proceedings of the National Academy of Sciences, 98(26), 15387–15392. https://doi.org/10.1073/pnas.251541498
Lei, Y., Liu, S., Guo, X., & Zuo, B. (2025). Neural synchronization and its impact on intergroup attitudes in dynamic interactions. Communications Biology, 8(1), 314. https://doi.org/10.1038/s42003-025-07776-8
Li, J., Wu, C., Li, Y., Pan, Y., Zhang, Y., Tian, S., Zhao, J., Wang, Y., & Liu, X. (2026). Oxytocin attenuates isolation-evoked emotional and social behavioural dysregulation through neural, immune and microbiota mechanisms. Translational Psychiatry, 16(1), 159. https://doi.org/10.1038/s41398-026-03888-9
Musardo, S., Contestabile, A., Knoop, M., Baud, O., & Bellone, C. (2022). Oxytocin neurons mediate the effect of social isolation via the VTA circuits. eLife, 11, e73421. https://doi.org/10.7554/eLife.73421
Nardou, R., Lewis, E. M., Rothhaas, R., Xu, R., Yang, A., Boyden, E., & Dölen, G. (2019). Oxytocin-dependent reopening of a social reward learning critical period with MDMA. Nature, 569(7754), 116–120. https://doi.org/10.1038/s41586-019-1075-9
Nardou, R., Sawyer, E., Song, Y. J., Wilkinson, M., Padovan-Hernandez, Y., de Deus, J. L., Wright, N., Lama, C., Faltin, S., Goff, L. A., Stein-O’Brien, G. L., & Dölen, G. (2023). Psychedelics reopen the social reward learning critical period. Nature, 618(7966), 790–798. https://doi.org/10.1038/s41586-023-06204-3
Puglia, M. H., Lynch, M. E., Nance, M. G., Connelly, J. J., & Morris, J. P. (2023). DNA methylation of the oxytocin receptor interacts with age to impact neural response to social stimuli. Frontiers in Aging Neuroscience, 15, 1252478. https://doi.org/10.3389/fnagi.2023.1252478
Rassovsky, Y., Harwood, A., Zagoory-Sharon, O., & Feldman, R. (2019). Martial arts increase oxytocin production. Scientific Reports, 9(1), 12980. https://doi.org/10.1038/s41598-019-49620-0
Sallet, J., Mars, R. B., Noonan, M. P., Andersson, J. L., O’Reilly, J. X., Jbabdi, S., Croxson, P. L., Jenkinson, M., Miller, K. L., & Rushworth, M. F. S. (2011). Social network size affects neural circuits in macaques. Science, 334(6056), 697–700. https://doi.org/10.1126/science.1210027
Schladt, T. M., Nordmann, G. C., Emilius, R., Kudielka, B. M., de Jong, T. R., & Neumann, I. D. (2017). Choir versus solo singing: Effects on mood, and salivary oxytocin and cortisol concentrations. Frontiers in Human Neuroscience, 11, 430. https://doi.org/10.3389/fnhum.2017.00430
Schwartz, L., Levy, J., Hayut, O., Netzer, O., Endevelt-Shapira, Y., & Feldman, R. (2024). Generation WhatsApp: Inter-brain synchrony during face-to-face and texting communication. Scientific Reports, 14(1), 2672. https://doi.org/10.1038/s41598-024-52587-2
Seeley, W. W., Menon, V., Schatzberg, A. F., Keller, J., Glover, G. H., Kenna, H., Reiss, A. L., & Greicius, M. D. (2007). Dissociable intrinsic connectivity networks for salience processing and executive control. Journal of Neuroscience, 27(9), 2349–2356. https://doi.org/10.1523/JNEUROSCI.5587-06.2007
Shao, L.-X., Liao, C., Gregg, I., Davoudian, P. A., Savalia, N. K., Delagarza, K., & Kwan, A. C. (2021). Psilocybin induces rapid and persistent growth of dendritic spines in frontal cortex in vivo. Neuron, 109(16), 2535–2544.e4. https://doi.org/10.1016/j.neuron.2021.06.008
Sherman, G. D., Rice, L. K., Jin, E. S., Jones, A. C., & Josephs, R. A. (2017). Sex differences in cortisol’s regulation of affiliative behavior. Hormones and Behavior, 92, 20–28. https://doi.org/10.1016/j.yhbeh.2016.12.005
Siegel, J. S., Subramanian, S., Perry, D., Kay, B. P., Gordon, E. M., Laumann, T. O., Reneau, T. R., Metcalf, N. V., Chacko, R. V., Gratton, C., Horan, C., Krimmel, S. R., Shimony, J. S., Schweiger, J. A., Wong, D. F., Bender, D. A., Scheidter, K. M., Whiting, F. I., Padawer-Curry, J. A., … Dosenbach, N. U. F. (2024). Psilocybin desynchronizes the human brain. Nature, 632(8023), 131–138. https://doi.org/10.1038/s41586-024-07624-5
Spreng, R. N., Dimas, E., Mwilambwe-Tshilobo, L., Dagher, A., Koellinger, P., Nave, G., Ong, A., Kernbach, J. M., Wiecki, T. V., Ge, T., Li, Y., Holmes, A. J., Yeo, B. T. T., Turner, G. R., Dunbar, R. I. M., & Bzdok, D. (2020). The default network of the human brain is associated with perceived social isolation. Nature Communications, 11(1), 6393. https://doi.org/10.1038/s41467-020-20039-w
Sumner, B. E. H., & Fink, G. (1998). Testosterone as well as estrogen increases serotonin 2A receptor mRNA and binding site densities in the male rat brain. Molecular Brain Research, 59(2), 205–214. https://doi.org/10.1016/S0169-328X(98)00148-X
Testard, C., Brent, L. J. N., Andersson, J., Chiou, K. L., Negron-Del Valle, J. E., DeCasien, A. R., Acevedo-Ithier, A., Stock, M. K., Antón, S. C., Gonzalez, O., Walker, C. S., Foxley, S., Compo, N. R., Bauman, S., Ruiz-Lambides, A. V., Martinez, M. I., Skene, J. H. P., Horvath, J. E., Cayo Biobank Research Unit, … Sallet, J. (2022). Social connections predict brain structure in a multidimensional free-ranging primate society. Science Advances, 8(15), eabl5794. https://doi.org/10.1126/sciadv.abl5794
Tomova, L., Wang, K. L., Thompson, T., Matthews, G. A., Takahashi, A., Tye, K. M., & Saxe, R. (2020). Acute social isolation evokes midbrain craving responses similar to hunger. Nature Neuroscience, 23(12), 1597–1605. https://doi.org/10.1038/s41593-020-00742-z
Wingfield, J. C., Hegner, R. E., Dufty, A. M., Jr., & Ball, G. F. (1990). The “challenge hypothesis”: Theoretical implications for patterns of testosterone secretion, mating systems, and breeding strategies. American Naturalist, 136(6), 829–846. https://doi.org/10.1086/285134
Yang, J., Zhang, H., Ni, J., De Dreu, C. K. W., & Ma, Y. (2020). Within-group synchronization in the prefrontal cortex associates with intergroup conflict. Nature Neuroscience, 23(6), 754–760. https://doi.org/10.1038/s41593-020-0630-x
Zhang, H., Yang, J., Ni, J., De Dreu, C. K. W., & Ma, Y. (2023). Leader–follower behavioural coordination and neural synchronization during intergroup conflict. Nature Human Behaviour, 7(11), 1972–1981. https://doi.org/10.1038/s41562-023-01663-0
Zink, C. F., Tong, Y., Chen, Q., Bassett, D. S., Stein, J. L., & Meyer-Lindenberg, A. (2008). Know your place: Neural processing of social hierarchy in humans. Neuron, 58(2), 273–283. https://doi.org/10.1016/j.neuron.2008.01.025